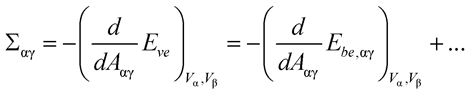 (1)
(1)Additions and corrections
Spontaneous tubulation of membranes and vesicles reveals membrane tension generated by spontaneous curvature
Reinhard Lipowskya
Faraday Discuss., 2013, 161, 305–331 (DOI: 10.1039/c2fd20105d). Amendment published 17th October 2013.
In section 6 of Ref.1, lipid vesicles enclosing two aqueous phases, α and β, have been considered. These phases typically form two droplets that partially wet the vesicle membrane and, thus, partition this membrane into two distinct membrane segments, αγ and βγ, with surface areas Aαγ and Aβγ. In Section 6.1, it was argued that the vesicle shapes involve only a single Lagrange multiplier tension, Σ, which is conjugate to the total membrane area A = Aαγ+Aβγ, because we have only a single constraint acting on this total area. The latter argument is, however, deceptive: in general, we need two Lagrange multiplier tensions Σαγ and Σβγ as one can see most clearly if one views these two quantities as the mechanical tensions that act to stretch (or compress) the two membrane segments.
Thus, consider the equilibrium shapes of vesicles with minimal energy Eve for fixed droplet volumes Vα and Vβ as a function of total membrane area A. For each value of A, these equilibrium shapes are characterized by certain segmental areas Aαγ and Aβγ. Likewise, the bending energy of these shapes consists of two terms, Ebe,αγ(Aαγ) and Ebe,βγ(Aβγ), corresponding to the two segments. The mechanical tensions Σαγ and Σβγ within these segments are then given by
(1)
and
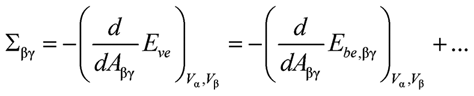 (2)
(2)
where the derivatives are taken at constant volumes Vα and Vβ. The dots indicate terms arising from additional energy contributions2 of the αβ interface and the contact line. The values for Σαγ and Σβγ as obtained from (1) and (2) will, in general, differ because the two membrane segments are exposed to different environments and are, thus, characterized by different elastic parameters. In addition, the segmental areas Aαγ and Aβγ can depend on the total membrane area A in a nonlinear manner, reflecting the variable partitioning of this area between the two segments.
For Σαγ ≠ Σβγ, the equations (66) and (67) in Ref.1, which describe the total membrane tensions 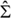 αγ and βγ, assume the form3
αγ and βγ, assume the form3
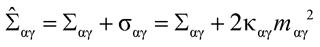 (66*)
(66*)
and
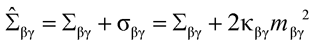 (67*)
(67*)
Furthermore, the relation (73) in Ref.1 is then replaced by
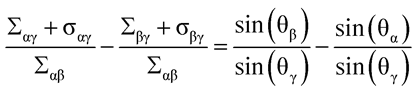 (73*)
(73*)
and the equations (77) - (79) are no longer valid. Finally, equation (80) in Ref.1 now attains the more general form
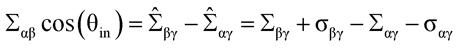 (80*)
(80*)
If the αγ membrane segment forms nanotubes, the mechanical tension Σαγ of this segment can be neglected because it is much smaller than the spontaneous tension σαγ,1 and the total tension
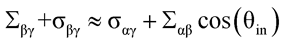 (3)
(3)
within the βγ segment must balance both the spontaneous tension σαγ of the αγ segment and the interfacial tension Σαβ of the liquid-liquid interface between the two droplets.
References
1 R. Lipowsky, Faraday Discuss., 2013, 161, 305–331.
2 H. Kusumaatmaja, Y. Li, R. Dimova and R. Lipowsky, Phys. Rev. Lett., 2009, 103, 238103.
3 Y. Li, R. Lipowsky and R. Dimova, Proc. Nat. Acad, Sci. USA, 2011, 108, 4731–4736.
a Theory and Bio-Systems, Max Planck Institute of Colloids and Interfaces, 14424 Potsdam, Germany;Fax: +49 331 567 9602; Tel: +49 331 567 9600; E-mail: lipowsky@mpikg.mpg.de
The Royal Society of Chemistry apologises for these errors and any consequent inconvenience to authors and readers.
Back to article